m6A去甲基化酶ALKBH5抑制胃癌细胞运动侵袭和Wnt信号通路活性的机
作者:admin 来源:未知 日期:2021-04-02 08:45人气:
摘 要:
目的 研究RNA去甲基化酶ALKBH5对胃癌细胞迁移和侵袭能力的影响及机制。方法 利用人类癌症基因库(the cancer genome Atlas, TCGA)对ALKBH5差异表达的胃癌患者进行总生存期分析;应用小鼠胃原位种植和转移模型分析ALKBH5的蛋白表达与胃癌细胞转移的相关性;通过构建敲低/过表达细胞株,结合Transwell实验研究ALKBH5对胃癌细胞迁移和侵袭能力的影响;利用m6A修饰位点预测网站m6AVAR(http://m6Avar.renlab.org/index.html),结合蛋白免疫印迹、放线菌素D处理结合实时荧光定量PCR,检测Wnt信号通路下游基因表达和mRNA稳定性。结果 TCGA数据库提示ALKBH5的低表达提示胃癌患者更短的生存期;体内实验验证ALKBH5的低表达与胃癌细胞远处转移相关;ALKBH5的敲低可以促进胃癌细胞的侵袭和转移;Wnt下游分子MYC,CD44,c-Met的mRNA上存在m6A修饰位点;ALKBH5缺失时,Wnt下游分子的蛋白表达增加及mRNA稳定性增强。结论 ALKBH5表达缺失促进胃癌细胞迁移和侵袭能力,维持Wnt信号通路的激活。
关键词:
ALKBH5 RNA甲基化 Wnt信号通路 胃癌侵袭
m6A demethylase ALKBH5 restrains migration and invasion and Wnt signaling pathway activity of gastric cancer cells
SHEN Liting LI Jinyang LI Xianfeng CHE Linrong LIU Kewei LIU Qin WANG Bin QIN Zhongyi CHEN Dongfeng
Department of Gastroenterology,Daping Hospital,Army Medical University (Third Military Medical University);
Abstract:
Objective To investigate the effect of RNA demethylase ALKBH5 on migration and invasion of gastric cancer(GC) cells and its potential mechanism. Methods Cancer Genome Atlas(TCGA) was employed to determine the correlation of ALKBH5 expression variations with overall survival(OS) in GC patients(n=371). The mouse models of orthotopic transplantation and metastasis were established to evaluate the correlation of between ALKBH5 expression and GC cells metastasis. After ALKBH5 knockdown and overexpression cells were constructed, the effect of ALKBH5 on migration and invasion of GC cells was explored with Transwell chamber assay. Furthermore, m6AVAR online database(http://m6Avar.renlab.org/index.html) was used to predict the m6A modification sites of Wnt activating targets; Western blotting and quantitative real-time PCR(qRT-PCR) combined with actinomycin D were applied to detect the protein expression and mRNA stability of downstream targets in Wnt signaling pathway. Results TCGA analysis showed that GC patients with lower ALKBH5 expression had shorter OS time. In vivo experiments confirmed that the low expression of ALKBH5 was involved in the distant metastasis of GC cells, and knockdown of ALKBH5 promoted the migration and invasion of GC cells. m6A modification sites were found in the mRNAs of Wnt downstream targets MYC, CD44 and c-Met, and the absence of ALKBH5 indicated increased protein expression and mRNA stability of these molecules. Conclusion The loss of ALKBH5 expression promotes the migration and invasiveness of GC cells and sustains the activation of Wnt signaling pathway.
Keyword:
ALKBH5; RNA methylation; Wnt signaling pathway; gastric cancer invasion;
胃癌(gastric cancer, GC)是我国最为常见的消化系统恶性肿瘤,肿瘤转移是胃癌患者不良预后的关键因素,究其原因是GC转移发生、发展的相关机制尚未完全阐明。表观稳态的失衡是维持肿瘤细胞恶性生物学行为的重要因素,RNA的动态化学修饰是表观调控的前沿领域。N6-甲基腺嘌呤N6-Methyladenosine(m6A)真核细胞中发现最早,丰度最高的RNA修饰方式[1-2]。m6A修饰参与基因表达调控中的多个环节,如RNA剪切,成熟,出核,翻译及降解,从而影响多种生理、病理进程,如胚胎发育,代谢稳态维持和肿瘤发生、发展[3]。m6A修饰酶的异常表达在胃肠道肿瘤发生、发展中的作用已有报道,但m6A是否参与胃癌转移尚不完全清楚。
m6A修饰的动态平衡由3大调控元件控制:编码器(Writer),主要指m6A转甲基转移酶复合物,包含核心催化酶METTL3(Methyltransferase-3),位点识别辅助蛋白METTL14(Methyltransferase-14),支架蛋白WTAP(Wilms tumor 1-associated protein)等;消码器(Eraser),m6A去甲基化酶,包括经典的FTO及ALKBH5(AlkB family member 5);读码器(Reader),主要由YTHDF1-3(YTH domain family1-3),YTHDC1/2(YTH domain-containing 1/2)和IGF2BPs构成。目前已有文献报道编码器和读码器在胃癌当中的作用,如METTL3通过m6A修饰调节HDGF(Hepatoma-derived growth factor)的表达进而维持胃癌细胞的糖酵解能力[4];YTHDF1通过提升FZD7(Frizzled7)翻译效率促进胃癌发生[5]。而消码器在胃癌中的作用仍未清楚阐述。FTO作为去甲基化酶不仅催化m6A去甲基化,还参与m6Am、m1A等去甲基化进程[6-7], ALKBH5相对于FTO,在底物的选择上更专一,仅催化去除m6A上的甲基,进而影响mRNA的剪接、稳定和翻译[8-9]。因此本研究重点探索ALKBH5作为m6A去甲基化酶在胃癌侵袭、转移中的作用及潜在调控机制。
Wnt信号通路是1条在生物进化过程中高度保守的通路,它参与调节多种细胞进程,如细胞增殖,分化,迁移以及干性维持。Wnt信号通路的异常激活可以促进一系列重要癌基因如MYC、c-Met的异常表达,进而驱动肿瘤的发生、发展[10-11]。有研究报道,METTL14的缺失可以通过抑制m6A修饰进而激活Wnt/PI3K-Akt信号,最终导致胃癌细胞的增殖以及侵袭[12]。但是在胃癌模型中,ALKBH5与Wnt信号通路之间的关系尚不清楚。本研究旨在探索m6A去甲基化酶ALKBH5是否通过刺激Wnt信号通路的激活参与到胃癌的恶性生物学行为的调控。
1 材料与方法
1.1 实验材料
1.1.1 主要试剂与仪器
包括:ALKBH5抗体,DDK标签抗体,HA标签抗体(日本MBL公司),FTO抗体, c-Met抗体,METTL3抗体,tubulin抗体,GAPDH抗体,IgG抗体(美国Proteintech公司),MYC抗体,β-catenin 抗体(美国Cell signaling technology公司),山羊抗鼠抗体,山羊抗兔抗体(北京中杉金桥生物技术公司),ALKBH5小发夹RNA,METTL3小发夹RNA,β-catenin质粒,METTL3质粒,ALKBH5质粒,GAPDH,MYC,CD44,c-Met PCR引物(上海生工生物工程有限公司),1640培养基,DMEM-高糖培养基,opti-MEM培养基,胎牛血清,Lipofectamine3000,青链霉素混合液(100×美国Gibco公司),Transwell小室(美国Corning公司),protein A /G-beads (美国Thermo公司),PCR反转录及定量扩增试剂(日本TAKARA公司),4%组织固定液,结晶紫染料,Western blot配胶试剂盒(上海碧云天公司),Western blot电泳槽,凝胶电泳成像仪(美国Bio-Rad公司),5% CO2细胞培养箱,酶标仪(美国Thermo公司)等。
1.1.2 细胞与动物
人正常胃黏膜上皮细胞GES-1,胃癌细胞(SGC-7901,MGC-803,BGC-823,HGC-27),人胚胎肾细胞HEK-293T购自中国科学院上海细胞库,胃癌细胞XNYY-0422由西南医院病理科赠送。BALB/C Nude小鼠由陆军特色医学中心实验动物中心提供。
1.2 方法
1.2.1 细胞培养
GES-1、SGC-7901、MGC-803、BGC-823、HGC-27细胞用含1%双抗,10%胎牛血清的1640培养基培养,HEK293T用含1%双抗、10%胎牛血清的DMEM高糖培养基培养。培养条件为:5% CO2,37 ℃恒温孵箱。每日观察,隔日换液,细胞汇合度85%~95%时传代。
1.2.2 包装病毒及慢病毒感染
将HEK-293T细胞传代至皿内,待细胞汇合度达到70%左右进行病毒包备,8 h后换液,24 h后收取上清,换入新鲜培养基,收取3次上清后浓缩病毒,准备病毒感染;将MGC-803,XNYY-0422传代至12孔板内,待细胞生长到50%左右行病毒感染,从-80 ℃冰箱取出浓缩的ALKBH5sh1、sh2、METTL3sh1、sh2病毒,过表达ALKBH5病毒和阴性对照病毒,冰上解冻。准备7只灭菌EP管,1管用于配置700含10%胎牛血清的DMEM高糖培养基与0.7 μL Polybrene混合液,分装至6只EP管内,各管90 μL,各管内分别加入10 μL ALKBH5 sh1、sh2、METTL3sh1、sh2病毒,过表达ALKBH5病毒和阴性对照病毒,混匀。弃12孔板内上清,各孔加入100 μL含病毒的混合液,孵箱培养,期间观察细胞形态,8 h后换入新鲜培养液,待汇合度达到80%~90%时传代至10 cm皿继续培养,细胞贴壁后行嘌呤霉素筛选。
1.2.3 过表达质粒转染
将HEK-293T细胞传代至6孔板内,待细胞汇合度达到70%左右时进行质粒转染。准备8只灭菌的EP管,8只管内均加入125 μL Opti-MEM ,其中4管内分别加入3、6、7.5、9 μL PEI,轻柔混匀,静置5 min; 另4只EP管内分别加入1 μg HA-β-catenin, 1 μg HA-β-catenin+1 μg Flag-METTL3,1 μg HA-β-catenin+1 μg Flag-METTL3+0.5 μg Flag-ALKBH5,1 μg HA-β-catenin+1 μg Flag-METTL3+1 μg Flag-ALKBH5轻柔混匀;将含有PEI的Opti-MEM培养基以前后对应的方式加入到包含质粒的混合液中,混合后静置10 min, 加入HEK-293T细胞内,孵箱培养,8 h后换液,待细胞汇合度95%时收集细胞部分提蛋白行Western blot实验,部分行免疫共沉淀实验。
1.2.4 Western blot实验
提取待实验细胞蛋白,检测蛋白表达水平。电泳恒压60 V/30 min, 120 V/1 h; 转膜:250 mA/1 h 10 min; 封闭:含5%奶粉的PBST溶液;一抗孵育4 ℃摇床过夜;二抗孵育室温摇床1 h; 曝光显影。
1.2.5 Transwell小室迁移与侵袭实验
共计4组细胞,MGC-803野生型细胞系,MGC-803敲低ALKBH5细胞系,XNYY-0422野生型细胞系,XNYY-0422过表达ALKBH5细胞系;四组细胞均饥饿处理24 h, 后消化,离心,PBS洗涤细胞1次去除残留血清,用无血清培养基重悬细胞,计数,MGC-803 1×104个/100 μL;XNYY-0422 1×104个/100 μL,加入到Transwell小室上室,下室加入700 μL含20%胎牛血清DMEM高糖培养基,迁移实验上下室中间聚碳酸酯膜无处理,侵袭实验聚碳酸酯膜上层铺基质胶。孵箱培养24 h后固定,染色,风干后显微镜下拍照。
1.2.6 实时荧光定量PCR(quantitative Real-time PCR,qRT-PCR)
利用TRIzol提取RNA后行反转录得到放线菌素处理后的MGC-803野生型和敲低ALKBH5细胞的cDNA产物,后以cDNA为模板扩增GAPDH、CD44、MYC和c-Met, qPCR程序为:95 ℃ 30 s; 95 ℃ 5 s, 60 ℃ 34 s, 72 ℃ 15 s, 共40个循环;95 ℃ 15 s, 60 ℃ 1 min, 95 ℃ 15 s。后根据c(T)值计算CD44,MYC和c-Met的相对表达量。
1.2.7 小鼠胃原位种植与转移模型
准备SGC-7901胃腺癌细胞系,使用PBS将细胞稀释至2×107个/mL。小鼠麻醉后行体表消毒,于腹部开1 cm小口,将小鼠胃轻柔取出,使用胰岛素针吸取50 μL细胞悬液,注入胃浆膜层下,注意防止细胞悬液从进针口渗出,体外观察无悬液渗漏后放回体内,缝合伤口。期间观察触摸是否形成肿瘤,2~3个月后处死小鼠,探查腹部脏器,拍照取证,取原发灶和转移灶胃癌细胞行下一步实验。
1.3 统计学分析
采用SPSS 20.0统计软件,数据采用x¯±s表示。TCGA数据库生存分析采用Log-rank检验,HR>1表示危险因素;Transwell小室迁移与侵袭实验采用独立样本t检验。检验水准α=0.05。
2 结果
2.1 ALKBH5低表达提示胃癌不良预后和远处转移
为明确ALKBH5是否与胃癌不良预后有关,我们通过人类癌症基因库(the cancer genome atlas, TCGA)对371例胃癌患者进行ALKBH5差异表达相关的总体生存期分析,发现ALKBH5低表达提示更短的生存期(HR=1.485,P<0.0340);与此同时,我们筛选出134例Ⅱ级胃癌和218例Ⅲ级胃癌进行生存分析,也得到相同的结果(HR=1.623,P=0.0861;HR=1.749,P=0.0163,图1)。通过构建胃癌原位种植和转移小鼠模型,获得原发灶及转移灶肿瘤细胞,提取蛋白进行Western blot检测,发现相对原发灶的胃癌细胞,转移灶中的FTO蛋白相对原发灶无明显差异,但ALKBH5蛋白表达水平更低(图1)。以上结果提示:①ALKBH5低表达与胃癌不良预后相关;②ALKBH5的低表达可能与胃癌细胞高侵袭转移特性相关。
2.2 ALKBH5敲低/过表达细胞系构建及验证
通过分析人正常胃黏膜细胞(GES-1),胃癌细胞株SGC-7901,MGC-803,BGC-823,HGC-27,XNYY-0422的ALKBH5的蛋白表达水平,发现在胃癌细胞MGC-803中,ALKBH5的表达水平相对较高,XNYY-0422中ALKBH5表达水平相对较低(图2A),因此我们分别选择MGC-803构建ALKBH5敲低细胞株,XNYY-0422构建过表达细胞株,通过Western blot验证2株敲低的MGC-803及1株过表达的XNYY-0422细胞株构建成功(图2B、C)。
2.3 ALKBH5的表达抑制胃癌细胞迁移和侵袭
应用Transwell实验观察敲低和过表达ALKBH5对胃癌细胞迁移和侵袭是否产生影响。结果提示:相对于对照组的MGC-803细胞,敲低ALKBH5的MGC-803细胞迁移和侵袭能力显著增强(P<0.000 1,P<0.000 1);在XNYY-0422细胞当中,过表达ALKBH5时细胞的迁移和侵袭能力减弱(P<0.000 1;P<0.000 1,图3)。以上结果表明:ALKBH5表达下调可以促进胃癌细胞的迁移和侵袭能力。
图1 ALKBH5表达降低与胃癌转移及不良预后密切相关
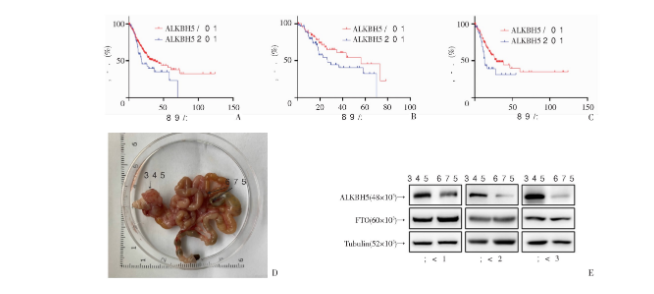
图2 ALKBH5敲低/过表达细胞系构建及验证

2.4 ALKBH5表达下调促进Wnt下游基因表达
Wnt信号通路的激活是维持胃癌高侵袭转移特性的关键分子生物学事件。ALKBH5作为经典的m6A去甲基化酶,是否影响Wnt信号通路的转导?通过比较对照组与敲低ALKBH5的MGC-803细胞系Wnt下游的靶基因表达情况,结果显示Wnt下游MYC与c-MET蛋白表达在敲低细胞系中表达增加(图4)。与此同时,METTL3作为公认的m6A核心甲基转移酶是否也参与Wnt信号通路的调节?我们通过构建敲低METTL3的MGC-803细胞系,发现当敲低METTL3时,Wnt下游MYC和c-MET表达出现抑制(图4)。以上结果提示:ALKBH5可能通过发挥其m6A消码器的作用参与调节Wnt信号通路下游基因表达。
图3 ALKBH5抑制胃癌细胞的运动侵袭能力 (结晶紫染色 ×200)
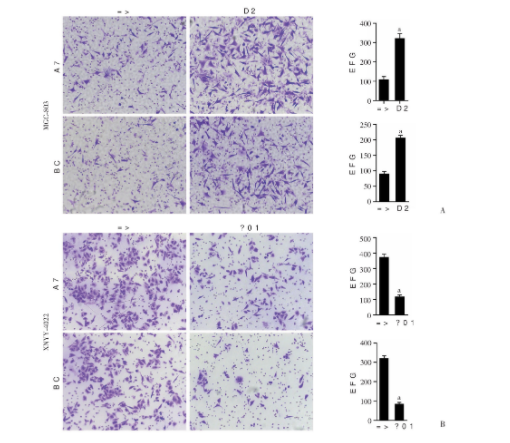
图4 ALKBH5和METTL3敲低Wnt信号下游基因表达的变化
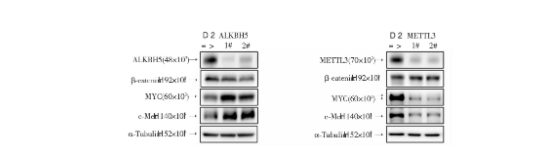
图5 MYC基因的m6A修饰位点及CD44和c-Met上潜在的m6A修饰位点
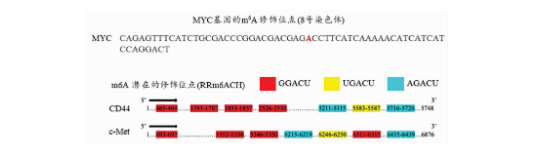
图6 放线菌素加入后MYC、CD44和c-Met mRNA含量的变化
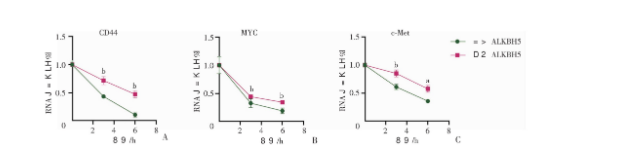
2.5 Wnt下游分子RNA m6A修饰位点预测
mRNA发生m6A修饰依赖于m6A修饰酶对底物的识别,那么Wnt信号通路下游的效应分子是否存在m6A修饰位点?我们运用m6A修饰位点预测网站m6AVAR(http: //m6Avar.renlab.org/index.html)发现了基于m6A交联免疫共沉淀法(m6A cross-linking immunoprecipitation, m6A-CLIP)测序数据中MYC基因的m6A修饰位点13,其具体位点见图5。同时基于m6A修饰位点的序列特征(RRACH,R=),我们发现CD44和c-Met上具备多个潜在的m6A修饰位点(图5)。以上结果提示Wnt下游关键效应分子MYC,CD44和c-Met可能受到RNA去甲基化酶ALKBH5的调节。m6A修饰通常通过影响mRNA稳定性参与基因表达调控,通过放线菌素抑制RNA合成结合qRT-PCR实验,我们发现敲低ALKBH5可显著延长MYC,CD44和c-Met的mRNA半衰期(图6)。上述结果表明ALKBH5可能通过去除Wnt下游靶基因MYC,CD44和c-Met mRNA上的m6A修饰维持其稳定性,促进基因表达。
3 讨论
胃癌是人类第5大常见肿瘤,也是最为常见的消化系统肿瘤,死亡病例数排第3位。胃癌起病隐匿,早期无特异的临床表现,大部分患者在首次确诊时已是中晚期,患者常常因此错过最佳治疗期[14]。因此,寻找到新的胃癌生物标志物和治疗靶点迫在眉睫。
m6A修饰已被证实在肿瘤发生和进展中起至关重要的作用,因此,m6A相关修饰酶是否可以作为肿瘤诊断标志物以及针对m6A修饰酶的抗肿瘤治疗策略是否有良好的临床运用前景亟须阐明。ALKBH5作为m6A去甲基化酶,已被证实在非小细胞肺癌[15],胶质瘤[16]、肝癌[17]、胰腺癌[18]发生和进展中起重要作用。本研究基于TCGA数据库行生信分析发现ALKBH5的低表达与胃癌不良预后相关,同时构建小鼠胃原位种植及转移模型证实ALKBH5低表达与胃癌远处转移相关,通过Transwell小室迁移与侵袭实验确认ALKBH5能够抑制胃癌细胞的侵袭转移,提示ALKBH5是胃癌进展中的抑癌因素。那么ALKBH5是如何参与调节胃癌进展的?文献[19]报道在胶质瘤中,ALKBH5可以去除SOX2转录子上的m6A修饰,上调SOX2的表达,激活Wnt信号,从而导致胶质瘤替莫唑胺(temozolomide, TMZ)耐药的发生;而在胰腺导管细胞癌中,ALKBH5可以通过m6A修饰Wnt抑制因子-1(Wnt inhibitory factor-1,WIF-1)的方式,激活WIF-1,下调Wnt信号,进而抑制胰腺癌细胞的增殖最终发挥其抑癌效应[20]。这些研究结果提示,ALKBH5表达失调,在不同肿瘤的恶性生物学行为中充当不同的角色,同时也说明ALKBH5-Wnt轴在肿瘤进展中发挥重要的调控作用。
在胃癌中,Wnt信号通路通常存在持续异常的激活并促进胃癌的进展[21]。文献[22-24]报道在多种类型肿瘤中,Wnt下游β-catenin、MYC、c-Met受到m6A修饰酶METTL3和IGF2BPs的调节。这些在胃癌中m6A修饰关键酶的过度激活或者缺失通常会介导Wnt信号通路下游基因表达异常。我们依托m6AVAR网站预测Wnt下游MYC、CD44和c-Met受m6A修饰的位点,同时基于mRNA半衰期实验,证明在胃癌细胞中,这些基因的表达也受到m6A去甲基化酶ALKBH5的调控。据此,我们的研究证明ALKBH5表达丢失介导的m6A修饰失衡可以导致Wnt信号下游分子的异常上调,可能是导致胃癌转移的潜在机制。结合胰腺癌中ALKBH5通过调控Wnt上游WIF-1表达进而发挥其抑癌效应,提示ALKBH5表达缺失导致的m6A修饰上调对于Wnt信号激活是明确的,m6A修饰的增加是促癌因素;但是,ZHANG等[12]报道在胃癌进展中,METTL14敲低介导m6A修饰的下调激活Wnt信号通路,提示m6A修饰的增加是抑癌因素。我们认为可能有以下原因:①m6A修饰水平的高低与编码器和消码器相关,但这种修饰发挥其转录调控的功能更多的基于读码器的作用;②ALKBH5缺失激活Wnt信号可能具备疾病依赖性,如何筛选出以m6A异常修饰作为主导调控方式的肿瘤亚型值得深入研究。
m6A修饰作为最常见的RNA修饰方式,广泛存在于转录产物mRNA上并影响其表达,是目前肿瘤表观靶向研究领域的热点。本研究基于临床实际问题,胃癌的高侵袭转移特性,发现m6A去甲基化酶ALKBH5表达的缺失可以显著促进胃癌细胞的迁移和侵袭能力;机制研究层面,我们初步探讨了ALKBH5可能通过去甲基化修饰Wnt信号通路下游关键基因MYC,CD44和c-Met, 促进其mRNA稳定性及蛋白表达。下一步我们将继续探索ALKBH5-Wnt轴的转录调控机制,同时明确读码器YTHDF1-3,YTHDC1/2在该调控轴中的作用,为探索胃癌转移的发生、发展机制以及通过靶向m6A的表观治疗策略,提供新的认识和研究思路。
参考文献
[1] MEYER K D,SALETORE Y,ZUMBO P,et al.Comprehensive analysis of mRNA methylation reveals enrichment in 3' UTRs and near stop codons[J].Cell,2012,149(7):1635-1646.DOI:10.1016/j.cell.2012.05.003.
[2] ROUNDTREE I A,EVANS M E,PAN T,et al.Dynamic RNA modifications in gene expression regulation[J].Cell,2017,169(7):1187-1200.DOI:10.1016/j.cell.2017.05.045.
[3] LEE Y,CHOE J,PARK O H,et al.Molecular mechanisms driving mRNA degradation by m6A modification[J].Trends Genet,2020,36(3):177-188.DOI:10.1016/j.tig.2019.12.007.
[4] WANG Q,CHEN C,DING Q Q,et al.METTL3-mediated m6A modification of HDGF mRNA promotes gastric cancer progression and has prognostic significance[J].Gut,2020,69(7):1193-1205.DOI:10.1136/gutjnl-2019-319639.
[5] PI J N,WANG W,JI M,et al.YTHDF1 promotes gastric carcinogenesis by controlling translation of FZD7[J].Cancer Res,2020:canres.0066.2020.DOI:10.1158/ 0008-5472.can-20-0066.
[6] MAUER J,LUO X B,BLANJOIE A,et al.Reversible methylation of m6Am in the 5’ cap controls mRNA stability[J].Nature,2017,541(7637):371-375.DOI:10.1038/nature21022.
[7] WEI J B,LIU F G,LU Z K,et al.Differential m6A,m6Am,and m1A demethylation mediated by fto in the cell nucleus and cytoplasm[J].Mol Cell,2018,71(6):973-985.e5.DOI:10.1016/j.molcel.2018.08.011.
[8] TANG C,KLUKOVICH R,PENG H Y,et al.ALKBH5-dependent m6A demethylation controls splicing and stability of long 3’-UTR mRNAs in male germ cells[J].Proc Natl Acad Sci USA,2018,115(2):E325-E333.DOI:10.1073/pnas.1717794115.
[9] ZHENG G Q,DAHL J A,NIU Y M,et al.ALKBH5 is a mammalian RNA demethylase that impacts RNA metabolism and mouse fertility[J].Mol Cell,2013,49(1):18-29.DOI:10.1016/j.molcel.2012.10.015.
[10] SCHOLZ B A,SUMIDA N,DE LIMA C D M,et al.WNT signaling and AHCTF1 promote oncogenic MYC expression through super-enhancer-mediated gene gating[J].Nat Genet,2019,51(12):1723-1731.DOI:10.1038/s41588-019-0535-3.
[11] HAN B,YAN S J,WEI S S,et al.YTHDF1-mediated translation amplifies Wnt-driven intestinal stemness[J].EMBO Rep,2020,21(4):e49229.DOI:10.15252/embr.201949229.
[12] ZHANG C,ZHANG M Q,GE S,et al.Reduced m6A modifi cation predicts malignant phenotypes and augmented Wnt/PI3K-Akt signaling in gastric cancer[J].Cancer Med,2019,8(10):4766-4781.DOI:10.1002/cam4.2360.
[13] KE S D,ALEMU E A,MERTENS C,et al.A majority of m6A residues are in the last exons,allowing the potential for 3’ UTR regulation[J].Genes Dev,2015,29(19):2037-2053.DOI:10.1101/gad.269415.115.
[14] RAWLA P,BARSOUK A.Epidemiology of gastric cancer:global trends,risk factors and prevention[J].Przeglad Gastroenterol,2019,14(1):26-38.DOI:10.5114/pg.2018.80001.
[15] JIN D,GUO J W,WU Y,et al.m6A demethylase ALKBH5 inhibits tumor growth and metastasis by reducing YTHDFs-mediated YAP expression and inhibiting miR-107/LATS2-mediated YAP activity in NSCLC[J].Mol Cancer,2020,19(1):40.DOI:10.1186/s12943-020-01161-1.
[16] ZHANG S C,ZHAO B S,ZHOU A D,et al.m6 A demethylase ALKBH5 maintains tumorigenicity of glioblastoma stem-like cells by sustaining FOXM1 expression and cell proliferation program[J].Cancer Cell,2017,31(4):591-606.e6.DOI:10.1016/j.ccell.2017.02.013.
[17] CHEN M N,WEI L,LAW C T,et al.RNA N6-methyladenosine methyltransferase-like 3 promotes liver cancer progression through YTHDF2-dependent posttranscriptional silencing of SOCS2[J].Hepatol Baltim Md,2018,67(6):2254-2270.DOI:10.1002/hep.29683.
[18] GUO X Y,LI K,JIANG W L,et al.RNA demethylase ALKBH5 prevents pancreatic cancer progression by posttranscriptional activation of PER1 in an m6A-YTHDF2-dependent manner[J].Mol Cancer,2020,19(1):91.DOI:10.1186/s12943-020-01158-w.
[19] LIU B Y,ZHOU J,WANG C Y,et al.LncRNA SOX2OT promotes temozolomide resistance by elevating SOX2 expression via ALKBH5-mediated epigenetic regulation in glioblastoma[J].Cell Death Dis,2020,11(5):384.DOI:10.1038/s41419-020-2540-y.
[20] TANG B,YANG Y H,KANG M,et al.m6A demethylase ALKBH5 inhibits pancreatic cancer tumorigenesis by decreasing WIF-1 RNA methylation and mediating Wnt signaling[J].Mol Cancer,2020,19(1):3.DOI:10.1186/s12943-019- 1128-6.
[21] 刘洪伟,梅基雄.双氢青蒿素通过Wnt/β-catenin信号通路抑制胃癌细胞的增殖和侵袭[J].第三军医大学学报,2018,40(5):387-394.DOI:10.16016/j.1000-5404.201710089.LIU H W,MEI J X.Dihydroartemisinin inhibits proliferation and invasion in human gastric cancer cells through suppression of Wnt/β-catenin signaling pathways[J].J Third Mil Med Univ,2018,40(5):387-394.DOI:10.16016/j.1000-5404.201710089.
[22] LIU L,WANG J,SUN G F,et al.m6A mRNA methylation regulates CTNNB1 to promote the proliferation of hepato blastoma[J].Mol Cancer,2019,18(1):188.DOI:10.1186/s12943-019-1119-7.
[23] HUANG H L,WENG H Y,SUN W J,et al.Recognition of RNA N6-methyladenosine by IGF2BP proteins enhances mRNA stability and translation[J].Nat Cell Biol,2018,20(3):285-295.DOI:10.1038/ s41556-018-0045-z.
[24] LUO G Y,XU W W,ZHAO Y P,et al.RNA m6A methylation regulates uveal melanoma cell proliferation,migration,and invasion by targeting c-Met[J].J Cell Physiol,2020,235(10):7107-7119.DOI:10.1002/jcp.29608.

