����֢ģ�ʹ�������С����ϸ����������֢���ӱ�������
���ߣ�admin ��Դ��δ֪ ���ڣ�2021-03-11 17:23������
ժ Ҫ��
Ŀ�� ̽�����Բ���Ԥ֪��Ӧ��(chronic unpredictable stress,CUS)����֢ģ�ʹ�����С����ϸ�����������С����ϸ������֢���ӵı仯��������� ѡȡ6��8���������Sprague-Dawley (SD)����,����Ӧ����������ˮƫ�û��ߵ�����ɸѡ����÷ֲ��������Ϊ�հ�����(n=19)��CUS��Ԥ��(n=48),����CUS��Ԥ�����5�ܵ�CUS��Ԥ,������ˮƫ��ʵ������������������������Ϊ,ɸѡ����ģ����16ֻ,��CUSģ���顣����������֯��ѧ������ѧ���ϵķ��������������������С����ϸ�������������о�ȷ�����о�,��������ӫ�ⷽ�������������������С����ϸ�����ܶ�,������ʵʱ����PCR������⺣����֢���ӵ�ˮƽ����� ��հ��������,CUSģ�������������������½�(P<0.01)����ˮƫ�ðٷֱ���������(P<0.01);CUSģ���������CA1��CA2/3��DG��С����ϸ�����������Ͽհ��������������(P<0.001,P=0.002,P=0.002);CUSģ���������CA1��CA2/3��DG���С����ϸ�����ܶȺͱ����Ͽհ��������������(P=0.011,P=0.015,P<0.001;P=0.018,P=0.019,P<0.001);����ڿհ�����,CUSģ�����������֢����IL-1β��iNOS��mRNA��������������(P=0.012,P=0.020)������ ����֢ģ�ʹ�������������С����ϸ�����������С����ϸ�����ܶȺͱ�������,��֢���ӱ�������,��������С����ϸ���ĸı���ܲ�������֢�IJ������̡�
�ؼ�����
����֢ ���� С����ϸ�� ��ϸ������1β �յ���һ��������ø ����ѧ
Increased number of microglia and enhanced expression of inflammatory factors in the hippocampus of rat model of depression
XIAO Kai LUO Yanmin LIANG Xin TANG Jing WANG Jin XIAO Qian QI Yingqiang LI Yue ZHU Peilin YANG Hao XIE Yuhan TANG Yong WU Hong
Department of Histology and Embryology,Laboratory of Stem Cells and Tissue Engineering,College of Basic Medical Sciences,Chongqing Medical University; Department of Physiology,College of Basic Medical Sciences,Chongqing Medical University; Department of Pathophysiology,College of Basic Medical Sciences,Chongqing Medical University; Department of Radioactive Medicine,College of Basic Medical Sciences,Chongqing Medical University;
Abstract��
Objective To investigate the changes of the total microglial number, density of activated microglia and inflammatory cytokines in the hippocampus of rat model of depression induced by chronic unpredictable stress(CUS). Methods Male Sprague-Dawley(SD) rats(6��8 weeks old) were randomly divided into control group(n=19) and CUS intervention group(n=48) with stratified randomization after adaptive feeding, sucrose preference baseline adjustment and screening. The rats of CUS intervention group were subjected to CUS intervention for 5 weeks. The depressive-like symptoms of 2 groups of rats were assessed with the sucrose preference test, and 16 model rats were screened out according to the results of sucrose preference. Immunohistochemical assay and stereological method were used to quantify the total number of microglia in the hippocampus of 2 groups of rats. The density of activated microglia in the hippocampus were studied with immunofluorescence assay. The contents of inflammatory cytokines in the hippocampus were measured with quantitative real-time polymerase chain reaction(qRT-PCR). Results The body mass and percentage of sucrose preference were significantly lower in the CUS model group than in the control group(P<0.01). The numbers of the microglia in the hippocampal CA1, CA2/3 and DG regions were significantly increased in the CUS model group compared with the control group(P<0.001, P=0.002, P=0.002), so were the density and proportion of the activated microglia in the CA1, CA2/3 and DG regions(P=0.011, P=0.015, P<0.001; P=0.018, P=0.019, P<0.001). Compared with the control group, the mRNA levels of inflammatory cytokines IL-1β and iNOS in the hippocampus were obviously higher in the model group(P=0.012, P=0.020). Conclusion The number of the microglia, density and proportion of activated microglia as well as levels of inflammatory cytokines are increased in the hippocampus of depression rats, indicating that the changes of hippocampal microglia might be involved in the pathogenesis of depression.
Keyword��
depression; hippocampus; microglia; interleukin-1beta; inducible nitric oxide synthase; stereology;
����֢��һ�ֳ����ľ���,���ٴ�֢״���������䡢���ȱʧ���ھθ�Ϊ��[1],���и߷����ʡ��߸����ʲ�������ɱ������ص�,���ؽ��ͻ��ߵ���������������ȫ��ļ�������[2]�����,����̽������֢�ķ������ơ�Ѱ���µ����ưе���зdz���Ҫ�����塣
��֢��Ӧ��Ϊ����֢��������Ҫ��˵֮һ[3],�ܵ�Խ��Խ�㷺�Ĺ�ע������������֢�����ı�Ĺؼ�����֮һ[4],ͬʱ,���о���������֢���߲�����֢��ػ�����ϵ��뺣������ļ����������[5]��С����ϸ����������ϵͳ�Ĺ�������ϸ��[6],�������е�С����ϸ��������֢����֢��Ӧ���������ŷdz���Ҫ������[7-8]�����о���������ӫ�⼼������������Ӧ���յ�������ģ��С���к���С����ϸ�����ܶ���������[9-10],�������Ƶ��о�����ͬ���ķ��������Բ���Ԥ֪��Ӧ��(chronic unpredictable stress,CUS)ģ��С��С����ϸ�������о�ȴ�ó����෴�Ľ���[11-12]�������о�����С����ϸ��������֢�ķ�������չ�������е���ϵ,��Ŀǰ����֢������С����ϸ���ı仯��Ȼ�������顣����,�����о���Ϊ�붨���о�,���о������ִ�����ѧ����[13]�Դ�������С����ϸ��������ά�����о�,���ڸ���ȷ��ϵͳ��̽������֢CUSģ�ʹ�������С����ϸ���ĸı䡣
С����ϸ���ڲ�ͬ������¾��о�Ϣ̬���̬�����ֲ�ͬ��״̬����Ӧ���Ȳ���������,С����ϸ���������ֳ��Ǩ�Ƶ���������,�������ܺ��������֢�ź�,����Ӱ����Ԫ��������ϸ��[14]��һ����Ӱ��ѧ�о�����,����֢���ߺ�������λ����(һ��С����ϸ������ı�־��)���ܶ���������[15]��ͬʱ,������Ӧ��ģ�Ͷ�����Ҳ����С����ϸ���Ļ����֢��������,���ָı��붯�������Ӧ�����µ���������Ϊ�������������[16]�����,�����ڻС����ϸ���Ͱ��������֢ʧ������������֢�ķ�չ����������������Ҫ������,������֢�������������С����ϸ���ľ���仯������д��о���
���о�����������֯��ѧ�����������ѧ���������о�����֢CUSģ�ʹ�������������С����ϸ�������ı仯,ͨ������ӫ�⼼��ϵͳ�о�CUSģ�ʹ�������������С����ϸ���Ļ����Լ�����ʵʱ����PCR�����۲��亣������֢�Ļ���������,���ڽ�һ����ʾ����֢�����IJ����ı�,Ϊ����֢�Ļ����о��ṩ��Ҫ�Ļ����о����ݡ�
1 �����뷽��
1.1 ����
1.1.1 ʵ�鶯��
ѡȡ6��8���䡢������Ϊ180��220 g������SD����(��������ҽ�ƴ�ѧʵ�鶯������),5ֻ/��,������ (22±1) �桢�㶨ʪ��(55±10)%����ҹ����Ļ�������Ӧ��ι��2��,���ɽ�ʳ��ˮ������ʵ�������ʵ�鶯������ͱ������йع涨���С�
1.1.2 �Լ���IJ�
�ÿ����ӻ��ƽ���νӷ���1(ionized calcium binding adapter molecule 1,Iba1)����(����ab178847)��С��CD68����(����ab955)����Ӣ��Abcam��˾;��SP�Լ���(����SP9001)��DAB ��ɫ�Լ���(����ZLI-9018)���Ա�����ɼ���Ź�˾;DyLight549�������IgGӫ�����(����A23320)��DyLight488�����С��IgGӫ�����(����A23210)��������Abbkine��˾�������Լ�:DAPIȾɫҺ(����AR1176,�人��ʿ�¹�˾);��RNA��ȡ�Լ���(����LS1040,�����������˾);cDNA��ת¼�Լ���(����RR047A,�ձ�TaKaRa��˾);ʵʱ����PCR�Լ���(����B21202,����Bimake��˾)��
1.1.3 ʵ������
����:������Ƭ��(CM1950,�¹�Leica��˾);����ѧ����ϵͳ(����Glostrup��˾);����ѧ����(BX51,�ձ�Olympus��˾);����۽�����(�¹�Zeiss��˾);ʵʱӫ�ⶨ��PCR��(CFX96,����Bio-Rad��˾)��
1.2 ����
1.2.1 ʵ�鶯����鼰CUSģ�͵Ľ���
�ھ���2�ܵ���ˮƫ�û��ߵ�����,������ˮƫ�ò��Խ��ɸѡ����ˮƫ�û���ƽ�ȵĴ���67ֻ, ���÷ֲ��������Ϊ�հ�����(n=19) �� CUS ��Ԥ��(n=48)���հ�������� (4��5ֻ/��) ����ι��,�������κ�Ӧ������������[17]�ķ�������CUSģ��,����Ӧ����Ԥ������ʳ/����(≤20 h)����ҹ�ߵ�(24��48 h)������(3��5 h)��ҹ��Ƶ��(8��12 h)����ƿ�̼�(2 h)����б(≥20 h)����Ϊ����(≥2 h)����ˮԡ(42 ��,5 min)����ˮԡ(4 ��,5 min)����ʪ����/�յ���(≥20 h)�����(1 min)��ҡ������β(1 min)�ȡ�CUS��Ԥ�������ι��,ÿ�����2��Ӧ��,����ҹ�ߵ���,ÿ��Ӧ����5 d�ڲ��ظ�,������ģ���̳���5�ܡ���Ӧ��������ÿ�̶ܹ�ʱ��Դ�������������ⶨ����ˮƫ�ò��ԡ���CUSӦ��������,������ˮƫ�ò��Խ��ɸѡ����ģ����16ֻ,��ΪCUSģ���顣
1.2.2 �������ⶨ
ÿ�̶ܹ�ʱ���������������������
1.2.3 ��ˮƫ��ʵ��
��ʵ������������֢ģ�Ͷ���ĺ���֢״——���ȱʧ�Ľ��[18]����ʵ�鿪ʼ֮ǰ,ÿֻ���������һ��ʱ�����ˮ��Ӧѵ��,ֱ���������ˮƫ�����ȶ������ŶԴ��������ˮƫ�ò���,���岽���������:���Ƚ�ʳ��ˮ24 h,Ȼ��ֱ����ÿֻ����1ƿ1%����ˮ��1ƿ��ˮ,��12 hʱ��������ƿ�ӵ�λ��,24 h��ֱ����ʣ����ˮ�Ͱ�ˮ��ƿ����,���ֱ�����Ӧ�������������������¹�ʽ������������ˮƫ�ó̶�:��ˮƫ�ðٷֱ�=��ˮ������/(��ˮ������+��ˮ������)×100%��
1.2.4 ��ʮ���Թ�ʵ��
�������۴��������컷���еĽ�������Ϊ[19]��װ�����������ۺ������ձ����,ÿ���۳�50 cm����10 cm����40 cm�����������װ�õ�����������,ͨ������ϵͳ��¼��5 min�ڴ���Ŀ��ۡ��ձ۽�������Լ��ڸ������ڵ�ͣ��ʱ��Ȳ�����
1.2.5 �걾ȡ�ĺ���Ƭ�Ʊ�
����Ϊѧ���Խ�����,ÿ�����ѡȡ5ֻ������ǻע��1%��ͱ�����(4 mL/kg)����������,�ʿ��ظ�ǻ����¶������,����ע�õ���ͷ�����ļⲢѸ�ټ������Ķ����ȹ�ע���и����Ƶ�������ˮ,��������ɫ��dzʱ������ע4%��ۼ�ȩֱ����֫��Ӳ����ע���������������Ĵ���,�������Ὣ�����з�Ϊ���Ұ���,����4%��ۼ�ȩҺ�й̶���ÿֻ�������ѡȡһ�����,���ݶ�������Һ(��0.1 mol/L��PBSϡ��)��ˮ,�����ٶ����ñ�����Ƭ�����Ŵ������״����50 μm�ĺ��������Ƭ������1/12�ij��������Ժ��к�����֯�IJ��ֽ��еȾ��������,ÿ�������õ�12����Ƭ������
1.2.6 ������֯��ѧȾɫ
��ÿ��ÿֻ�������Ƭ���������ѡȡ2����Ƭ,����Ƭ��������Ϊ1/6��������Ƭ��0.01 mol/L��PBSƯϴ10 min×3��,��0.3% Triton��Һ(0.3 mL Triton×100+0.1 mL Tween20+100 mL 0.01 mol/L PBS)Ưϴ10 min×6��;3%��Դ�Թ�������ܹ���20 min,0.3%Triton��ҺƯϴ15 min×2��;����������Һ��ˮԡ��30 min,0.3%Triton��ҺƯϴ10 min×2 ��;���Һ(10%ɽ��Ѫ�� SP9001��1%̥ţѪ�塢0.1% ��������0.3 mL Triton×100)37 ����2 h��Ȼ����뿹Iba1����4 �� ���� 48 h,0.3% Triton��ҺƯϴ10 min×6��;����(�����ر������IgG)37 �����3 h,0.3% Triton��ҺƯϴ10 min×6��;����(����ø�����ù�Ѱ���)37 �����2 h,0.3% Triton��ҺƯϴ10 min×6�Ρ�Ȼ��DAB�ܹ���ɫ1 min��ȥ����ˮ��PBS��Ưϴ10 min,�����Ƭ�̶�����ľ����Ⱦ���ݶ��Ҵ���ˮ�����ױ�����������֬��Ƭ��
1.2.7 ����ѧ��������
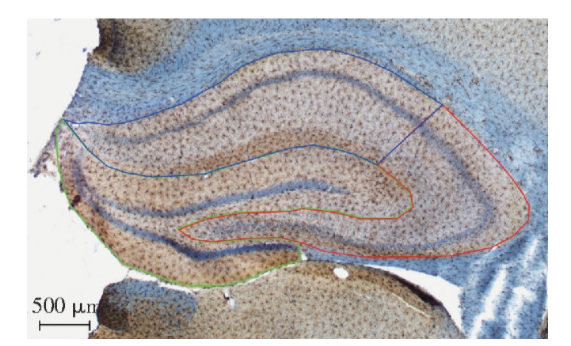
������ѧϵͳ�а��չ�ѧ�ֺϷ�[20]��Iba1+С����ϸ�����м���,���岽������:�������ݴ������ͼ�Ĺ�����2.5���ᄉ�»��ֺ���3�������ı߽�(ͼ1),Ȼ����100���;���ͳһ���ò���:���ӿ�ĸ߶�Ϊ15 μm,�ϱ��汣���߶�Ϊ3 μm,�����������Ϊ6 %��Ȼ����ݹ�ѧ���ӿ�Ľ��߷���(ͼ2),����ÿ����Ұ���Ը߶��ڰ��������۽���С����ϸ����Ŀ,ÿ��2����Ұ������Ƭ�ĺ��(��Ƭ�ϱ������±���֮��ľ���)��������ù�ѧ�ֺϷ����㺣����������Iba1+С����ϸ��������Ŀ,��ʽ����:
N=∑Q-×1/ssf×1/asf×1/tsf
∑Q-Ϊÿֻ������������������Iba1+ϸ��������;ssfΪ��Ƭ��������1/6;asfΪ�����������6%;tsf�Ǹ߶ȳ�������(���Կ�ĸ߶�/��Ƭ��ƽ�����)��
ͼ1 ������CA1��CA2/3��DG���ı߽�ʾ��ͼ
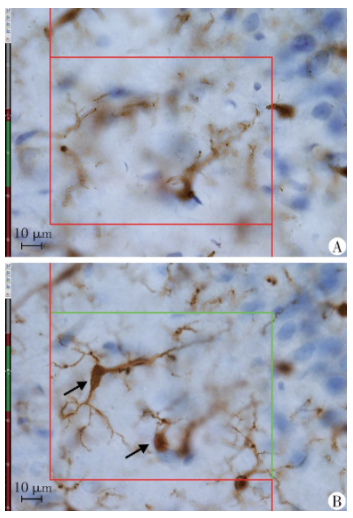
ͼ2 ���ӿ��������Iba1+С����ϸ��ʾ��ͼ
1.2.8 ����ӫ��Ⱦɫ��ϸ������
��ÿ��ÿֻ�������Ƭ���������ѡȡ1����Ƭ����Ƭ��PBSƯϴ10 min×3��, 0.3%Triton��Һ(0.3 mL Triton×100+0.1 mL Tween20+100 mL 0.01 mol/L PBS)Ưϴ20 min×3��;����������Һ��ˮԡ��30 min,0.3%Triton��ҺƯϴ10 min×2 ��;���Һ(10%ɽ��Ѫ�� SP-9001��1%̥ţѪ�塢0.3 mL Triton ×100)37 ����2 h�������뿹Iba1����Ϳ�CD68����,4 ��,����60 h,0.3% Triton��ҺƯϴ20 min×3��;����DyLight549�������IgGӫ�����(��ɫ)��DyLight488�����С��IgGӫ�����(��ɫ),37 �����2 h,PBSƯϴ5 min×6��;����DAPIȾϸ����,���·���5 min;��Ƭ,��Ƭ��ʹ�ü���۽�������200 �����������Ҫ���ӫ��ͼ��ÿֻ������Ƭ������ÿ���������ٻ��10�ž��д����Ե�ͼ��ÿ��ͼ���Iba1+/CD68+ϸ�������ֶ���������������ϸ�����ܶȡ�
1.2.9 ʵʱ����PCR
��ÿ�鶯�������ѡȡ5ֻ�������ʵʱ����PCR�������������������Ͼ���ȡ����������,Ѹ�ٷ����������֯,��Һ���䶳����-80 �汣�档����˵����IJ���,����RNA��ȡ�Լ��дӺ�������ȡ��RNA,�ڲⶨRNAŨ�Ⱥ�,����ת¼�Լ��н�����ת¼�õ�cDNA��IL-1β��������:5′-TGAG-GCTGACAGACCCCAAAAGAT-3′,��������:5′-GCTCC-ACGGGCAAGACATAGGTAG-3′,���ﳤ��181 bp;iNOS��������:5′-GAGACGCACAGGCAGAGGTTG-3′,��������:5′-AGCAGGCACACGCAATGATGG-3′,���ﳤ��123 bp;IL-6��������:5′-ACTTCCAGCCAGTTGCCTT-CTTG-3′,��������:5′-TGGTCTGTTGTGGGTGGTAT-CCTC-3′,���ﳤ��110 bp;TNF-α��������:5′-TCG-TAGCAAACCACCAAGCG-3′,��������:5′-AGAGAAC-GGATGAACACGCCA-3′,���ﳤ��452 bp;β-actin��������:5′-TGTCACCAACTGGGACGATA-3′,��������:5′-GGGGTGTTGAAGGTCTCAAA-3′,���ﳤ��165 bp��ʹ��ʵʱ����PCR�Լ��н���PCR������Ӧ,����ʵʱӫ�ⶨ��PCR�Ǽ��PCR�IJ�������β-actinΪ�ڲ�,��2-ΔΔCt���Ի�������mRNA����������ͳ�Ʒ�����
1.3 ͳ��ѧ����
ʹ��SPSS 23.0ͳ�����������ݽ��з����������Ƚ�����̬�Լ����,��������̬�ֲ�������x¯±s��ʾ,��������λ��(�ķ�λ�����)��ʾ�����������ݲ����ظ������ķ������,��ʮ���Թ�ʵ�鲿�����ݲ���Mann-Whitney U����,�������ݵ������Ƚ�����ö�������t���顣����ˮα=0.05��
2 ���
2.1 CUSģ�ʹ��������������
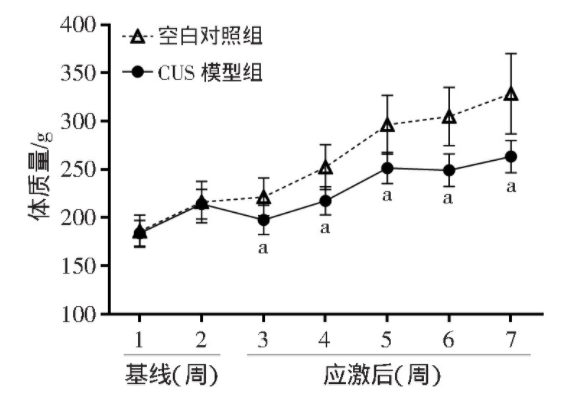
�ڻ��ߵ�����,CUSģ���������հ���������������������ͳ��ѧ���塣CUS��Ԥ��ʼ��,�հ���������������������,CUSģ��������������������ٶȽϿհ����������仺,��CUSģ����������������հ����������������(P<0.01,ͼ3)��
ͼ3 �������CUS��Ԥǰ���������仯
2.2 CUSģ�ʹ������ˮƫ�ðٷֱȽ���
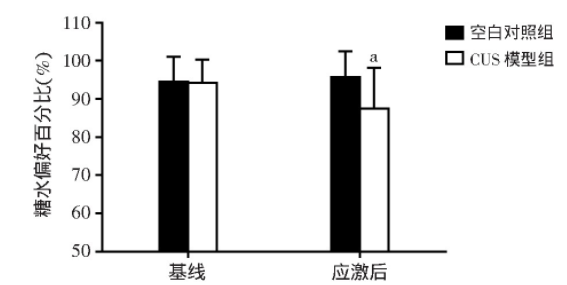
��Ӧ����Ԥǰ,CUSģ����Ϳհ����������ˮƫ�ðٷֱȲ�����ͳ��ѧ����(t=1.058,P=0.298);�ڳ���5�ܵ�Ӧ����Ԥ��,CUSģ����������ˮƫ�ðٷֱȽϿհ����������������(t=4.488,P<0.01,ͼ4)��
2.3 CUSģ�ʹ����н�������Ϊ
CUSģ�������Ϳհ��������Ľ����ܴ��������۽��������ͣ��ʱ�䡢�ձ۽��������ͣ��ʱ�䡢���۽�������ٷֱ��Լ�����ͣ��ʱ��ٷֱȵȲ����������ͳ��ѧ����(��1)��
ͼ4 CUS��Ԥǰ�����������ˮƫ�ðٷֱ�
2.4 CUSģ�ʹ�������С����ϸ������������
ͼ5Ϊ���������������Iba1+С����ϸ����֯��ѧȾɫ�Ĵ�����ͼ��,��հ����������������С����ϸ������̬���,CUSģ�����Iba1+С����ϸ���ķֲ���Ϊ�ܼ�����������ѧ���������������,CUSģ���������CA1��Iba1+С����ϸ�������Ͽհ�������������(t=-8.141,P<0.001), CA2/3��Iba1+С����ϸ�������Ͽհ�������������(t=-4.376,P=0.002), DG��Iba1+С����ϸ�������Ͽհ�������������(t=-4.427,P=0.002,ͼ6)��
��1 ��������ʮ���Թ�ʵ����[x¯±s,��λ��(�ķ�λ�����)]
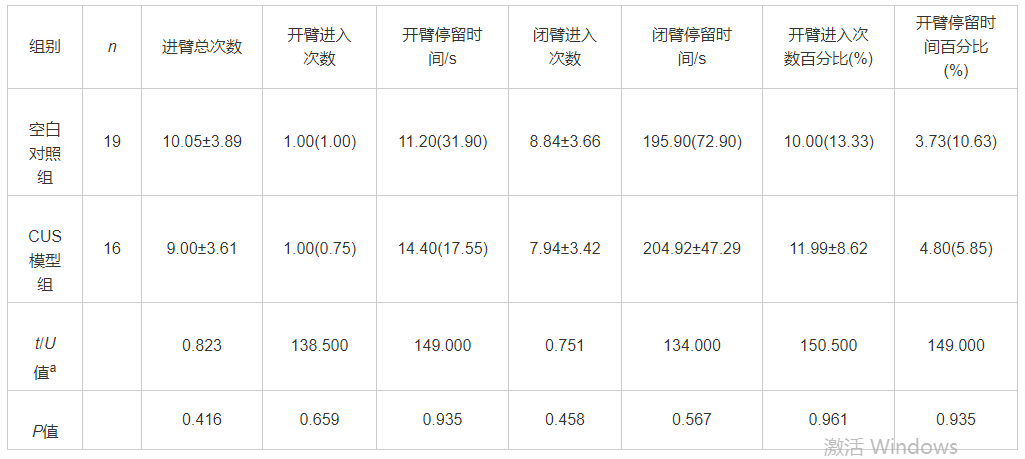
ͼ5 �������������Iba1+С����ϸ���������黯Ⱦɫ�۲� (S-P)
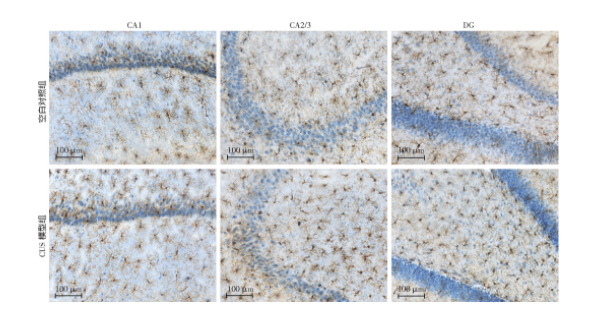
ͼ6 �������������Iba1+С����ϸ���������ıȽ�
2.5 CUSģ�ʹ������ڻС����ϸ���ܶȺͱ�������
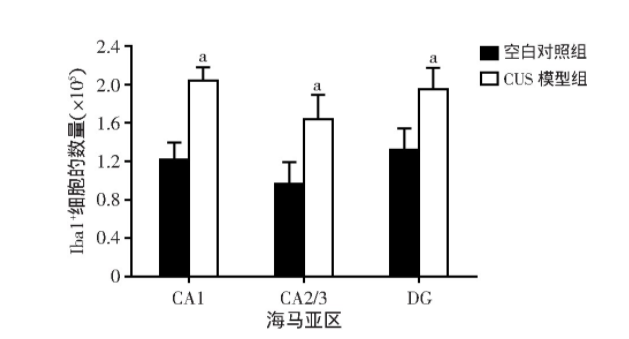
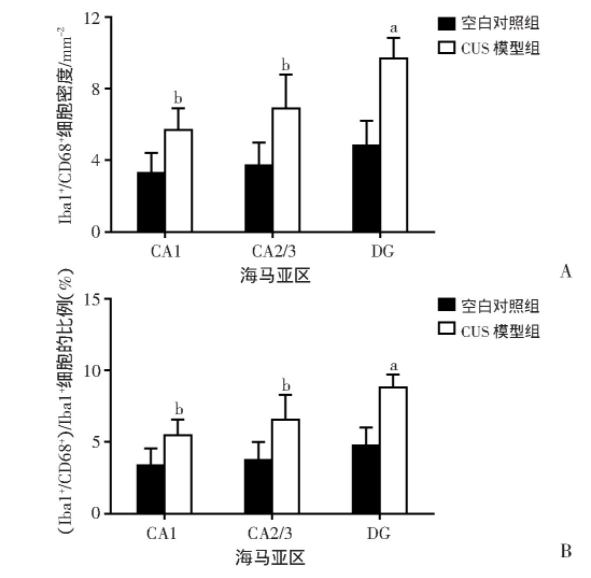
ͼ7Ϊ������Iba1+/CD68+С����ϸ������ӫ��Ⱦɫ�Ĵ�����ͼ��,��հ��������,CUSģ�������������Iba1+/CD68+С����ϸ���ֲ���Ϊ�ܼ���ͨ����������:��հ��������,CUSģ���������CA1��Iba1+/CD68+С����ϸ�����ܶ���������(t=-3.271,P=0.011);��հ��������,CUSģ���������CA2/3��Iba1+/CD68+С����ϸ�����ܶ���������(t=-3.099,P=0.015);��հ��������,CUSģ���������DG��Iba1+/CD68+С����ϸ�����ܶ���������(t=-6.094,P<0.001)��ͬʱ,��հ��������,CUSģ���������CA1��Iba1+/CD68+С����ϸ��ռIba1+С����ϸ���ı�����������(t=-2.958,P=0.018);��հ��������,CUSģ���������CA2/3��Iba1+/CD68+С����ϸ��ռIba1+С����ϸ���ı�����������(t=-2.913,P=0.019);��հ��������,CUSģ���������DG��Iba1+/CD68+С����ϸ��ռIba1+С����ϸ���ı�����������(t=-5.801,P<0.001,ͼ8)��
ͼ7 �������С����ϸ��������ӫ��Ⱦɫ�۲�
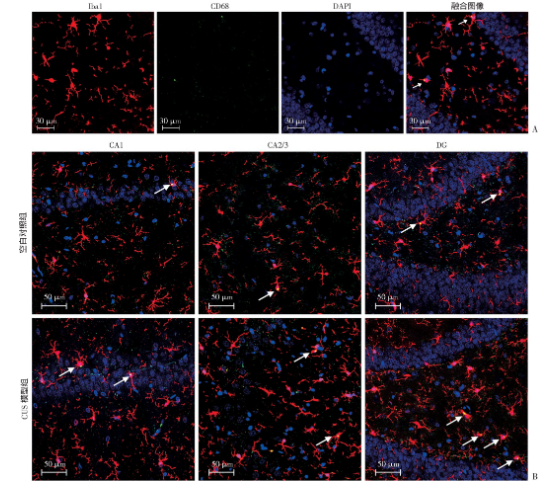
ͼ8 ��������������С����ϸ������
2.6 CUSģ�ʹ�������֢����IL-1β��iNOS��mRNA����ˮƽ����
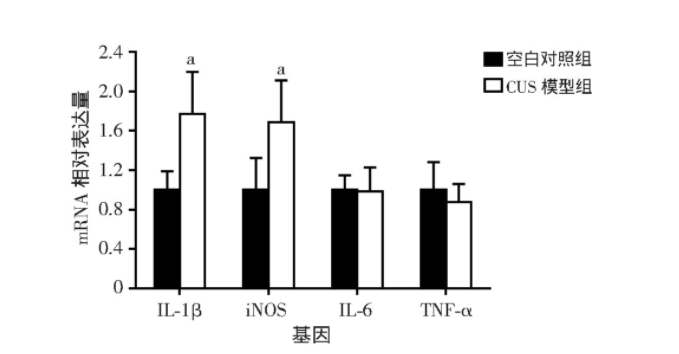
ͨ��ʵʱ����PCR��ⷢ��,CUSģ�����������֢����IL-1β��iNOS��mRNA�������Ͽհ�������������(t=-3.667,P=0.012;t=-2.884,P=0.020),��CUSģ����Ϳհ������������֢����IL-6��TNF-α��mRNA������������ͳ��ѧ����(ͼ9)��
ͼ9 ʵʱ����PCR������������IL-1β��iNOS��IL-6�� TNF-α mRNA����
3 ����
����֢��һ�ֳ���������������뻷���ȶ�����ص��µľ��������ϰ��Լ���,����������Ӧ��������֢����Ҫ�²�����֮һ[21]�����о����þ����CUS���յ��������������Ϊ,��������ˮƫ��ʵ������������֢����ĺ���֢״——���ȱʧ���������:CUS��Ԥ��Ĵ�����ˮƫ�ðٷֱ���������,�������������������ٶ����Լ��������Ͻ��������Ϊ��5�ܵ�CUS��Ԥ������֢����ģ�ͽ����ɹ�������,���о�Ҳ���ø�ʮ���Թ�ʵ����������������������Ϊ,�����ʾCUSģ����Ϳհ�����ĸ����ʮ���Թ������������ͳ��ѧ����,������ʵ�������֢CUSģ�ʹ����н��ǡ�
���������Ӧ���߶�����[22],�������о�ָ�����������Dz�������֢����֢��Ӧ����Ҫ����֮һ[5]��Ŀǰ�������������֢���о��������ں�����Ԫ�Ĺ����ϰ���,�Խ���ϸ����С����ϸ�����о����١�С����ϸ���������ڵ��˾���ϸ������������ЧӦϸ��[6],��Ӧ���Ȳ��������С����ϸ�����Ա��������Ԫ����������[14]��Iba1������С����ϸ��(������Ϣ̬�ͻ̬)������Ա���[23],��ĿǰС����ϸ���ȽϹ��ϵı�����С����ϸ��Iba1�����ķ���[24]�������֢״����������������Ԥ������֢�Ľ��̡��ڱ�ʵ����,���Dz��þ�ȷ������ѧ������������CUSģ�ʹ�����CA1��CA2/3��DG��Iba1+С����ϸ�����������������ӡ���ǰ���о���������ӫ�⼼��Ҳ����������Ӧ���յ�������ģ�Ͷ�������������DG����С����ϸ�����ܶ���������[9-10],����Ҳ���о���������ӫ�⼼����������ģ��С������С����ϸ���ܶ���������[11-12]����Щ��������ӫ�⼼�����о���ʹ�ð붨���ķ����Ժ�����άƽ���С����ϸ����������,����С����ϸ���������ı仯���п���ʵ�ķ�ӳ�����о�������ƫ����ѧ������ά��ȷ����������������С����ϸ���ı仯,������Ӿ�ȷ�ɿ���ȫ�档���о������������ģ�ʹ�����С����ϸ������Ŀ�����ӿ���������֢����Ҫ�����ı�֮һ��
����С����ϸ�����������仯��,С����ϸ���Ļ������֢�ķ�����չ������Ҳ����ʮ����Ҫ������[25]��CD68�ǻС����ϸ������������[26],��ˮƽ������֢ģ������������[27]��FENG��[28]���о����������º�Ӧ��(chronic mild stress,CMS)������С����DG���/����С����ϸ�����ܶȡ�Ȼ������CUS����ģ�ʹ������������ڻС����ϸ���ľ���仯���д��о������о�ͨ����Iba1����Ϳ�CD68���������ӫ��Ⱦɫ����ǻ״̬��С����ϸ������ϼ���۽������۲�,����CUS���������˴�����CA1��CA2/3��DG��Iba1+/CD68+ϸ�����ܶȼ�ռIba1+С����ϸ���ı���,��������Ӧ���յ��˺�����������С����ϸ���Ļ���ڲ���������,С����ϸ������ƫ����̬��ת��Ϊ����״̬[14]��ͬʱ,����Ӧ����ص�����(��Ӧ���������ߡ�����ˮƽ�ı��)�������շ�С����ϸ���ļ���[29]���ڱ��о���,CUS��Ԥ�����ĸ���Ӧ�������ǻС����ϸ�����ӵ���Ҫ����֮һ��Ȼ��,��Ӧ����ϵ���е�С����ϸ������ľ�������д���һ���о���
����,���о�������CUS��Ԥ�����˴�������֢����IL-1β��iNOS�Ļ������ˮƽ,��Щ�������ǰ���о�һ��[30-31]��Ȼ������֢ģ�ʹ�����IL-6��TNF-α�Ļ������ˮƽû�������仯,�Ʋ����������Ϊ����֢����֢��Ӧ�����ܵ�������ص�Ӱ�졣ͬʱ,С����ϸ�����ڵ�IL-1β��iNOS��������������֢����֢��Ӧ��������Ӧ�IJ����ı�[32-33],��˵������֢������֢������IL-1β��iNOS��IL-6��TNF-α�ı仯����������֮,���о��������С����ϸ���Ļ����֢����IL-1β��iNOS�����߿��ܴٽ�����֢�ķ�������չ��
��������,���о�������ƫ����ѧ����������ӫ�⼼����ʵʱ����PCR����������֢ģ�ʹ�������С����ϸ��������֢���ӽ������о�,�����ʾ����ģ�ʹ���������С����ϸ�����������ӡ��С����ϸ���ܶȺͱ��������Լ���֢��������,��ʾ����С����ϸ������������֢�����ı����Ҫ�ṹ������δ�������о�����֢С����ϸ���ı�ľ������,���л���Ϊ����֢�ṩ�µ����ưе�����ƴ�ʩ��
�����
[1] BARNETT R.Depression[J].Lancet,2019,393(10186):2113.DOI:10.1016/S0140-6736(19)31151-1.
[2] OTTE C,GOLD S M,PENNINX B W,et al.Major depressive disorder[J].Nat Rev Dis Primers,2016,2:16065.DOI:10.1038/nrdp.2016.65.
[3] MILLER A H,RAISON C L.The role of inflammation in depression:from evolutionary imperative to modern treatment target[J].Nat Rev Immunol,2016,16(1):22-34.DOI:10.1038/nri.2015.5.
[4] TAYLOR W D,MCQUOID D R,PAYNE M E,et al.Hippocampus atrophy and the longitudinal course of late-life depression[J].Am J Geriatr Psychiatry,2014,22(12):1504-1512.DOI:10.1016/j.jagp.2013.11.004.
[5] MAHAJAN G J,VALLENDER E J,GARRETT M R,et al.Altered neuro-inflammatory gene expression in hippocampus in major depressive disorder[J].Prog Neuropsychopharmacol Biol Psychiatry,2018,82:177-186.DOI:10.1016/j.pnpbp.2017.11.017.
[6] WAKE H,MOORHOUSE A J,MIYAMOTO A,et al.Microglia:actively surveying and shaping neuronal circuit structure and function[J].Trends Neurosci,2013,36(4):209-217.DOI:10.1016/j.tins.2012.11.007.
[7] TORRES-PLATAS S G,CRUCEANU C,CHEN G G,et al.Evidence for increased microglial priming and macrophage recruitment in the dorsal anterior cingulate white matter of depressed suicides [J].Brain Behav Immun,2014,42:50-59.DOI:10.1016/j.bbi.2014.05.007.
[8] CALCIA M A,BONSALL D R,BLOOMFIELD P S,et al.Stress and neuroinflammation:a systematic review of the effects of stress on microglia and the implications for mental illness [J].Psychopharmacology (Berl),2016,233(9):1637-1650.DOI:10.1007/s00213-016-4218-9.
[9] DONG S Q,ZHANG Q P,ZHU J X,et al.Gypenosides reverses depressive behavior via inhibiting hippocampal neuroinflammation[J].Biomed Pharmacother,2018,106:1153-1160.DOI:10.1016/j.biopha.2018.07.040.
[10] ZHAO Q,WU X,YAN S,et al.The antidepressant-like effects of pioglitazone in a chronic mild stress mouse model are associated with PPARγ-mediated alteration of microglial activation phenotypes[J].J Neuroinflam,2016,13(1):259.DOI:10.1186/s12974-016-0728-y.
[11] CAI Z,YE T,XU X,et al.Antidepressive properties of microglial stimulation in a mouse model of depression induced by chronic unpredictable stress[J].Prog Neuropsychopharmacol Biol Psychiatry,2020,101:109931.DOI:10.1016/j.pnpbp.2020.109931.
[12] TONG L,GONG Y,WANG P,et al.Microglia loss contributes to the development of major depression induced by different types of chronic stresses[J].Neurochem Res,2017,42(10):2698-2711.DOI:10.1007/s11064-017-2270-4.
[13] WEST M J,GUNDERSEN H J.Unbiased stereological estimation of the number of neurons in the human Hippocampus[J].J Comp Neurol,1990,296(1):1-22.DOI:10.1002/cne.902960102.
[14] YIRMIYA R,RIMMERMAN N,RESHEF R.Depression as a microglial disease[J].Trends Neurosci,2015,38(10):637-658.DOI:10.1016/j.tins.2015.08.001.
[15] SETIAWAN E,WILSON A A,MIZRAHI R,et al.Role of translocator protein density,a marker of neuroinflammation,in the brain during major depressive episodes [J].JAMA Psychiatry,2015,72(3):268-275.DOI:10.1001/jamapsychiatry.2014.2427.
[16] DAI J,DING Z,ZHANG J,et al.Minocycline relieves depressive-like behaviors in rats with bone cancer pain by inhibiting microglia activation in Hippocampus[J].Anesth Analg,2019,129(6):1733-1741.DOI:10.1213/ane.0000000000004063.
[17] WILLNER P,TOWELL A,SAMPSON D,et al.Reduction of sucrose preference by chronic unpredictable mild stress,and its restoration by a tricyclic antidepressant[J].Psychopharmacology,1987,93(3):358-364.DOI:10.1007/BF00187257.
[18] WILLNER P,MUSCAT R,PAPP M.Chronic mild stress-induced anhedonia:a realistic animal model of depression[J].Neurosci Biobehav Rev,1992,16(4):525-534.DOI:10.1016/s0149-7634(05)80194-0.
[19] WALF A A,FRYE C A.The use of the elevated plus maze as an assay of anxiety-related behavior in rodents[J].Nat Protoc,2007,2(2):322-328.DOI:10.1038/nprot.2007.44.
[20] TANG Y,JANSSEN W G M,HAO J D,et al.Estrogen replacement increases spinophilin-immunoreactive spine number in the prefrontal cortex of female rhesus monkeys[J].Cereb Cortex,2004,14(2):215-223.DOI:10.1093/cercor/bhg121.
[21] HAMMEN C.Stress and depression[J].Ann Rev Clin Psych,2005,1:293-319.DOI:10.1146/annurev.clinpsy.1.102803.143938.
[22] THOMAS R M,HOTSENPILLER G,PETERSON D A.Acute psychosocial stress reduces cell survival in adult hippocampal neurogenesis without altering proliferation [J].J Neurosci,2007,27(11):2734-2743.DOI:10.1523/JNEUROSCI.3849-06.2007.
[23] ITO D,TANAKA K,SUZUKI S,et al.Enhanced expression of Iba1,ionized calcium-binding adapter molecule 1,after transient focal cerebral ischemia in rat brain[J].Stroke,2001,32(5):1208-1215.DOI:10.1161/01.str.32.5.1208.
[24] SINGHAL G,BAUNE B T.Microglia:an interface between the loss of neuroplasticity and depression[J].Front Cell Neurosci,2017,11:270.DOI:10.3389/fncel.2017.00270.
[25] WACHHOLZ S,ESSLINGER M,PLUMPER J,et al.Microglia activation is associated with IFN-alpha induced depressive-like behavior[J].Brain Behav Immun,2016,55:105-113.DOI:10.1016/j.bbi.2015.09.016.
[26] MURPHY J E,TEDBURY P R,HOMER-VANNIASINKAM S,et al.Biochemistry and cell biology of mammalian scavenger receptors[J].Atherosclerosis,2005,182(1):1-15.DOI:10.1016/j.atherosclerosis.2005.03.036.
[27] XU N,TANG X H,PAN W,et al.Spared nerve injury increases the expression of microglia M1 markers in the prefrontal cortex of rats and provokes depression-like behaviors[J].Front Neurosci,2017,11:209.DOI:10.3389/fnins.2017.00209.
[28] FENG X,FAN Y,CHUNG C Y.Mefenamic acid can attenuate depressive symptoms by suppressing microglia activation induced upon chronic stress[J].Brain Res,2020,1740:146846.DOI:10.1016/j.brainres.2020.146846.
[29] WOHLEB E S,FRANKLIN T,IWATA M,et al.Integrating neuroimmune systems in the neurobiology of depression[J].Nat Rev Neurosci,2016,17(8):497-511.DOI:10.1038/nrn.2016.69.
[30] LIU L L,LI J M,SU W J,et al.Sex differences in depressive-like behaviour may relate to imbalance of microglia activation in the hippocampus[J].Brain Behav Immun,2019,81:188-197.DOI:10.1016/j.bbi.2019.06.012.
[31] ���h,���ڷ�,�δ�÷,��.Sirt1ͨ���ٽ�������С����ϸ����M2��ת�����������ºͲ���Ԥ֪��Ӧ��ģ��С�����������Ϊ[J].������ҽ��ѧѧ��,2019,41(14):1301-1307.DOI:10.16016/j.1000-5404.201901077.WANG Y,WAN T F,DUAN C M,et al.Sirt1 ameliorates CUMS-induced depression-like behaviors in the mice via promoting hippocampal microglia M2-polarization[J].J Third Mil Med Univ,2019,41(14):1301-1307.DOI:10.16016/j.1000-5404.201901077.
[32] BROWN G C,NEHER J J.Microglial phagocytosis of live neurons[J].Nat Rev Neurosci,2014,15(4):209-216.DOI:10.1038/nrn3710.
[33] HE M C,SHI Z,SHA N N,et al.Paricalcitol alleviates lipopolysaccharide-induced depressive-like behavior by suppressing hypothalamic microglia activation and neuroinflammation[J].Biochem Pharmacol,2019,163:1-8.DOI:10.1016/j.bcp.2019.01.021.

